



Overview of Equine Arboviral Encephalomyelitis
(Equine viral encephalitis)
- Equine Arboviral Encephalomyelitis
- Overview of Equine Arboviral Encephalomyelitis
Equine encephalitides can be clinically similar, usually cause diffuse encephalomyelitis (see Meningitis, Encephalitis, and Encephalomyelitis) and meningoencephalomyelitis, and are characterized by signs of CNS dysfunction and moderate to high mortality. Arboviruses are the most common cause of equine encephalitis, but rabies virus, Sarcocystis neurona (see Equine Protozoal Myeloencephalitis), Neospora hughesii (see Neosporosis), equine herpesviruses, and several bacteria and nematodes may also cause encephalitis. Arboviruses are transmitted by mosquitoes or other hematophagous insects, infect a variety of vertebrate hosts (including people), and may cause serious disease. Most pathogenic arboviruses use a mosquito to bird or rodent cycle. Tickborne encephalitides are also a differential cause in the eastern hemisphere. Arboviral diseases are ever emerging, and there are arboviruses pathogenic to horses on virtually every continent.
Etiology and Epidemiology of Arboviral Encephalitides:
Alphaviruses:
North America is home to some of the most pathogenic encephalitic viruses because of the enduring endemic status of alphaviruses of the family Togaviridae. Endemic species in North, Central, or South America include Eastern equine encephalitis virus (EEEV), Western equine encephalitis virus (WEEV), Highlands J virus, Venezuelan equine encephalitis virus (VEEV), Everglades virus, and Una virus (see Table: Arboviruses That Can Cause or Are Linked to Equine Encephalomyelitis). Other alphaviruses associated with equine encephalitis are Semliki Forest virus in Africa and Ross River virus in Australia and the South Pacific. EEEV has two distinct antigenic variants that are separated longitudinally. The North American variant is the most pathogenic and is found in eastern Canada; all states within the USA east of the Mississippi River and in Arkansas, Minnesota, South Dakota, and Texas; and in the Caribbean Islands. The South American variant is less pathogenic and confined to central and South America and is now called Madariaga virus (MADV).
Arboviruses That Can Cause or Are Linked to Equine Encephalomyelitis
EEE and WEE viruses are separated in North America primarily latitudinally; however, WEEV is relatively heterogeneous, with several subtypes consisting of WEE, Highlands J virus, Sindbis, Aura, Ft. Morgan, and Y 62–33. Several variants have been found in horses (see Table: Arboviruses That Can Cause or Are Linked to Equine Encephalomyelitis). WEE is found in western Canada, states in the USA west of the Mississippi, and in Mexico and South America. WEE previously isolated in the south and eastern USA has been shown to belong to the Highlands J virus serogroup. VEE has six antigenically related subtypes: subtype I, Everglades, Mucambo, Pixuna, Cabassou, and AG80–663. Subtype I serovars AB and C primarily cause epizootics; subtype I and serovar IE caused a large outbreak in Mexico in 1993. Epizootic strains are not generally found in the USA, although there was an epizootic of VEE in 1971. Sylvatic subtype II (Everglades) has been isolated from people and mosquitoes in Florida; subtype III has been isolated in the Rocky Mountains and northern plains states.
The general life cycle of alphaviruses involves transmission between birds and/or rodents and mosquitoes. In North America, theoretically, EEEV is perpetuated in a sylvatic cycle between avian hosts (passerine birds) and mosquitoes, with primary transmission in this cycle via Culiseta melanura. Birds do not develop disease but develop sufficient viremia for transmission to mosquitoes unless the bird belongs to an introduced species that in North America includes European starlings, house sparrows, pheasants, and emus. Field work in Alabama indicates that the northern cardinal is the primary target for C melanura feeding; however, other mosquitoes (C erraticus) are capable of feeding on a wider variety of birds, including robins, chickadees, owls, and mockingbirds. Mammalian reservoirs may also be important during years of high EEEV transmission. Experimental infection of cotton rats, a marshland rat found throughout the Americas, resulted in viremia just being capable of transmitting EEEV, with juvenile rats developing higher titers and 100% mortality to infection.
Horses and people are clinically affected but do not develop viremia sufficiently high enough to transmit virus to mosquitoes and so are considered “dead-end” hosts. However, young animals are more susceptible to EEEV, and it is not known how important non-neural tissues are for harboring high viral infection. Presumed extraneural sites of infection include cardiac tissue and bone marrow. During epizootics, alpacas, llamas, cattle, swine, cats, and dogs can develop disease. Snakes have been identified as a possible reservoir.
Freshwater hardwood swamps are the most associated enzootic niche for EEEV. In the southern USA, which has the highest number of reported annual cases, reemergence within mammalian hosts is associated with “tree farms” that often function as inland freshwater swamps. Nonetheless, EEEV disease occurs frequently in naive horses in southeastern habitats not associated with sylvatic field ecology, so a full understanding of the epizootic cycle still remains to be elucidated. Intense focal activity has been reported in Michigan, Wisconsin, Ohio, Massachusetts, and New Hampshire. In 2005, Massachusetts experienced a human case affected rate more than five times that of the preceding 10 yr; affected people resided within ½ mile of a cranberry bog or swamp associated with forest habitat.
In Central and South America, the principal vectors of MADV belong to Culex (Melanoconion) spp. These vectors feed on birds, rodents, marsupials, and reptiles, with rodent reservoirs possibly featuring more importantly in this life cycle. Before the year 2000, comparatively few epizootics of disease caused by MADV in horses were recorded in South America, with minimal disease reported in people. Furthermore, there are notable differences in virulence between EEEV strains in South America vs in North America. In 2008 and 2009, larger outbreaks occurred in Central and South America. In northeastern Brazil, 229 horses were affected, with a case fatality rate of 73% and disease severity similar to that of EEEV in North America.
WEE is transmitted primarily by C tarsalis, which is found just west of the Mississippi river and throughout the West. This mosquito breeds in sunlit marshes and in pools of irrigation water in pastures. WEE can also be transmitted by the tick Dermacentor andersoni. Epizootics of WEE are associated with increased rainfall in early spring followed by warmer than normal temperatures.
Sylvatic VEE viruses are found throughout North, Central, and South America in jungle or swampy areas, Two cycles occur with VEEV, the enzootic cycle and the epizootic cycle. The mosquitoes that serve as the primary vectors for the bird- or rodent-mosquito life cycle are members of the subgenus Melanoconion (C cedecci). Epizootics are associated with a mutation to a subtype I (AB, C, and possibly E), a change in mammalian pathogenesis, and a change to several bridge vectors. The enzootic cycle centers around sylvatic rodents such as spiny and cotton rats, which have high natural infection rates and can develop viremia high enough to transmit VEEV to mosquitoes. Opossums, bats, and shore birds likely contribute to dispersal of the enzootic virus so that constant cycling occurs. When epizootics occur with mutation of the virus and this change in mosquito vectors, equine infection becomes a predominant feature in the maintenance of epizootic VEE. All mammalian hosts are capable of developing a high-titer viremia of ~106 plaque-forming units/mL for up to 5 days, but the horse is likely to be the most important mammalian host in terms of vector capacity.
Flaviviruses:
In general, viruses belonging to the Flaviviridae and Bunyaviridae families are less pathogenic than the Togaviridae; however, viral encephalomyelitis caused by any of these pathogens is a potentially catastrophic illness for any vertebrate host. There are 53 species of flavivirus, and many are clinical pathogens for horses, including Japanese encephalitis, West Nile virus (WNV) encephalitis, Kunjin virus (KUN), and Murray Valley encephalitis virus. Overall, the diseases caused by the Japanese encephalitis serogroup are similar (see Table: Arboviruses That Can Cause or Are Linked to Equine Encephalomyelitis). All of these viruses are transmitted by a mosquito vector, with Culex spp usually the most efficient transmitter. KUN is actually a strain of WNV and is found in Australia, some southeast Asia countries, and New Guinea. Disease in horses caused by Murray Valley fever is geographically restricted to the South Pacific and is sporadic in occurrence.
WNV has the widest geographic distribution of all of the flaviviruses. Prior to 1999, WNV was recognized in Africa, the Middle East, Asia, and occasionally in European countries. In 1999, WNV infection was first recognized in North America. Since then, the virus has spread throughout the USA and parts of Canada and Mexico. WNV isolated from the outbreak in New York in 1999 appears to be closely related to an isolate recovered from geese in Israel in 1998.
Since 2010, continued worldwide spread and reemergence of many flaviviruses, especially WNV, has occurred. As of 2014, WNV is recognized as having seven lineages, with lineage 1 (sublineage 1a, 1b, and 1c) and lineage 2 affecting people and horses. Lineage 1a activity in the USA and Europe dominated much of the end of the 1990s until the mid-2000s, causing neuroinvasive infections in people and horses in Africa, Europe, Australia, Asia, North and Central America, and the Middle East. In 2012, a very large epizootic occurred in the USA, with 5,674 human and 690 equine cases reported. New emergences have occurred in Greece and Serbia, and the first lineage 1a case in India was detected in 2012. Lineage 1b viruses are primarily represented by KUN, with Australia undergoing the largest epizootic of arboviral disease in 2011, involving 900 horses. Although multiple pathogens were detected, including KUN, Murray Valley encephalitis virus, and Ross River virus, there was emergence of a new strain of KUN. The 1c viruses are fairly nonpathogenic and found in India. (This sublineage may soon be designated as lineage 5.) Lineage 2 strains, long considered African based and inducing mild disease in horses, is also expanding globally with increased neuroinvasive activity. Neuroinvasive disease due to lineage 2 has emerged across Europe in Hungary, Austria, Italy, and Greece, which had at least four epizootic cycles with cases of neuroinvasive disease identified in horses.
Both wetland and terrestrial birds may be involved in the natural cycle of WNV, with migratory birds thought to introduce the virus into a geographic region. However, a wide range of infected birds (~326) have high, sustained viremia and little or no clinical disease (passerines). Fatal infections among corvids (eg, crows, blue jays, and magpies) have been the hallmark of WNV infection in the USA. Ticks have been demonstrated to be infected with WNV, but their role in natural transmission is unknown. Experimentally, transmission has been documented between cohabitating birds and from oral exposure to WNV in drinking water in birds. Oral transmission has been demonstrated experimentally in several types of raptors.
In people, other important routes of infection include blood transfusions, organ donation, breast milk, and across the placenta. Sporadic infections and illness have also occurred in several other mammalian species, including dogs, cats, camelids, sheep, and squirrels. Oral transmission has been demonstrated experimentally in cats. Farmed alligators have demonstrated disease and mortality due to WNV, and there has also been a report of WNV-induced disease in crocodiles. Alligators are susceptible to oral infection. In addition to birds, only alligators have consistently demonstrated high enough viremia (104–105 plaque-forming units) to amplify virus, serve as reservoir hosts, and transmit virus back to mosquitoes.
Most equine-associated flaviviruses are maintained in an enzootic transmission cycle between wild birds and mosquitoes, although tickborne encephalitis found in Eurasia can cause disease in horses. Many species of mosquitoes can transmit the equine virulent viruses, although Culex spp are principal vectors to maintain enzootic activity. C pipiens complex and C tarsalis are thought to play the largest role in natural transmission in North America. In the eastern and Midwestern regions of the USA, C pipiens is one of the major vectors, whereas in the western regions of the USA, C tarsalis is thought to be one of the most efficient vectors.
Bunyaviruses:
Cache Valley virus (transmitted by mosquitoes and Culicoides sp among rabbits), Main Drain virus (transmitted by Culicoides varipennis among hares and rodents in the western USA), and snowshoe hare virus (transmitted by Culiseta and Aedes mosquitoes among rabbits in southern Canada and northern USA) have all been identified, although infrequently, as the cause of encephalitis in horses.
Clinical Findings of Viral Encephalomyelitis in Equids:
The initial clinical signs are similar for the arboviruses; progression of clinical signs and severity of disease are the differentiating features. Initially, horses are quiet and depressed, with clinical neurologic signs generally occurring 9–11 days after infection. Compared with WEE and WNV disease, clinical signs of EEEV (and VEEV) encephalomyelitis more frequently include altered mentation, impaired vision, aimless wandering, head pressing, circling, inability to swallow, irregular ataxic gait, paresis and paralysis, seizures, and death. Spinal signs are often symmetric with ataxia in all limbs, rapidly progressing to quadriparesis, along with intensification of forebrain signs. Many horses progress to recumbence within 12–18 hr of onset of neurologic abnormalities. Most deaths occur within 2–3 days after the onset of signs.
The clinical signs and course of disease are highly variable in WNV disease and other flavivirus encephalomyelitis. Presenting complaints most often include neurologic abnormalities; other common initial complaints include colic, lameness, anorexia, and fever. Initial systemic signs include a mild fever, feed refusal, and depression. Neurologic signs are highly variable, but spinal cord disease and moderate mental aberrations are most consistent. Spinal cord disease manifests as asymmetric, multifocal or diffuse ataxia and paresis. Severe manifestations of WNV may occur independently in the fore- or hindlimbs, unilaterally, or in a single limb. In all clinical studies published to date, for WNV, >90% of affected horses developed some type of spinal cord signs, whereas 40%–60% developed behavioral changes characterized by periods of hyperesthesia, ranging from mild apprehension to overt hyperexcitability, with fractious reactions to aural, visual, and tactile stimuli. Fine and coarse tremors of the face and neck muscles are common, described in 60%–90% of horses. Some horses have periods of cataplexy or narcolepsy that may render them temporarily or permanently recumbent.
Cranial nerve deficits can be seen in all arbovirus infections of horses; these include most cranial nerves with cell bodies located in the mid- and hindbrains. Weakness and/or paralysis of the face and tongue are most frequent. Horses with facial and tongue paresis can be dysphagic, and overt signs of quidding or even esophageal choke can develop. Many horses with severe mental depression and facial paresis will keep their heads low, resulting in severe facial edema. Occasionally, head tilt may be seen. Infrequently in WNV disease, urinary dysfunction ranging from mild straining to stranguria has been reported, making differentiation from equine herpesvirus 1 (EHV-1) more challenging.
During the neurologic phase, horses frequently thrash and injure themselves. Sepsis from trauma in recumbent horses also occurs. Prolonged recumbency leads to pulmonary infections, especially in foals, in which a long duration of slinging and treatment may be pursued more frequently than in large, recumbent animals. Dysphagia leads to decreased water and food intake, with renal damage due to concurrent use of anti-inflammatory drugs. Skin and muscle necrosis are common in recumbent horses. Life-threatening trauma can also occur, including a ruptured diaphragm and fractures.
Lesions:
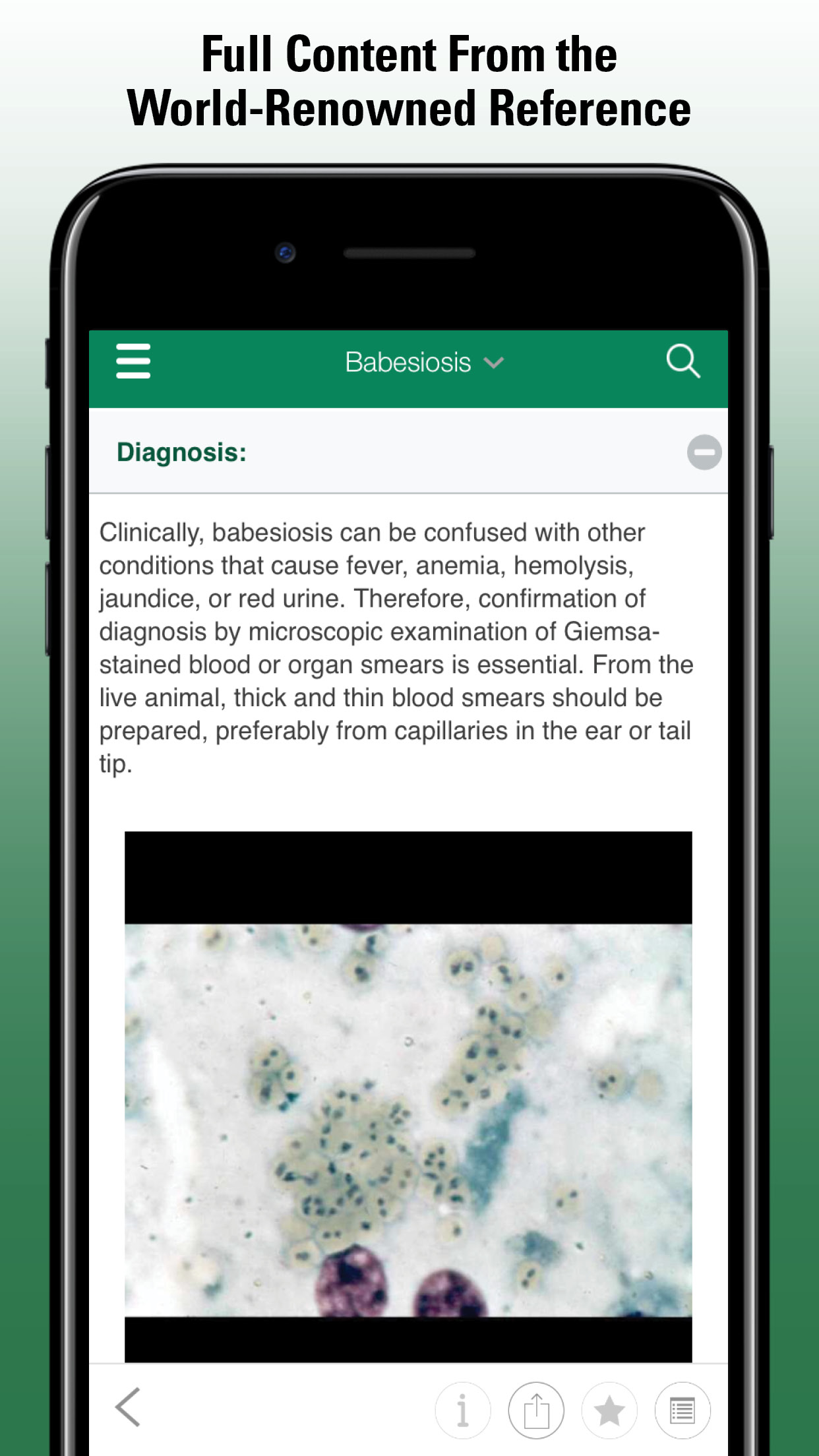
All necropsies on horses suspected of viral encephalitis should be performed with all participants wearing personal protective gear (ie, waterproof gown, boots, gloves, N-95 or N-99 mask, face shield, and hair covering). Gross lesions are most common with EEEV and severe VEEV disease and are characterized by widespread and prominent congestion of the meninges. In other infections, gross lesions are rarer and are limited to small multifocal areas of discoloration and hemorrhage throughout the brain and spinal cord. The brain should be examined microscopically for the presence of meningoencephalitis. In EEEV infection, a severe gliosis with necrosis of the neuropil is seen in the cerebrum and extending through the corona radiata through the hindbrain and cervical spinal cord. Although mononuclear cells are present, neutrophils are widespread and diffusely distributed. The gliosis is less nodular than in WNV, and when there are accumulations of cells, there is extensive necrosis and often frank microscopic hemorrhage within and around lesions. In milder cases and in WNV infections, microscopically there is a non-necrotizing lymphohistiocytic poliomeningoencephalitis. Slight to severe inflammation, characterized by perivascular cuffing of lymphocytes and monocytes, is usually present. In more severe cases, there can be extravasation of fluid and red blood cells from vessels. Often the distribution of lesions is multifocal, with more severe lesions present in grey matter. In the neuropil, dying neurons often are surrounded by microglial cells. The most severe gliosis and perivascular cuffing is often in the midbrain (thalamus, hypothalamus) and hindbrain (pons and medulla), extending into the cerebellum. Lesions can be multifocal in the spinal cord and can be more severe in the lumbar cord.
Diagnosis:
No consistent changes in clinical pathology have been found in equine viral encephalitis. With EEEV and WNV in horses, peripheral lymphopenia is common. Horses are frequently azotemic, likely from decreased food and water intake.
In general, no pathognomonic signs distinguish flavivirus infection in horses from other CNS diseases, and a full diagnostic evaluation should be performed. Confirmation of flavivirus infection with encephalitis in horses begins with assessment of 1) whether the horse meets the case definition based on clinical signs; (2) whether the horse resides in an area in which flavivirus has been confirmed in the current calendar year in mosquitoes, birds, people, or horses; and (3) lack of appropriate vaccination.
In terms of antemortem diagnostic testing, analysis of CSF can be a valuable adjunct to presumptive clinical assessment. CSF analyzed from horses with acute EEEV infection typically shows a neutrophilic pleocytosis with markedly increased total solids. Horses infected with EEEV that are partially immune may have predominantly mononuclear cells, but nondegenerate neutrophils are still present. Although WNV-infected horses can have normal CSF, if the CSF is abnormal, there is a mononuclear pleocytosis with moderately to markedly increased total solids. In a few horses with acute infections, virus may be isolated from the CSF. By the time neurologic signs are seen, viremia has ended and detection of virus in the plasma of clinically affected horses is of no value.
Serology is the key to antemortem diagnosis of recent alphavirus and flavivirus infection in horses showing clinical signs. IgM antibody rises sharply and is increased in 85%–90% of horses with clinical arboviral encephalitis. Thus, the IgM capture ELISA is the test of choice to detect recent exposure to these viruses. Neutralizing antibody titers (primarily IgG) develop slowly during this time and stay increased for several months. Although neutralizing antibody tests will differentiate between subtypes of these viruses, and are thus considered the gold standard for confirmatory serology, paired serum samples are essential to detect recent exposure and to differentiate antibody response due to field infection from vaccine responses for any of these viruses. Because virus-neutralizing antibodies appear at the end of viremia and may precede appearance of neurologic signs, paired samples may not show a 4-fold increase in horses while demonstrating neurologic signs. In horses that succumb without premortem and postmortem testing, paired samples from febrile herdmates may be necessary to confirm presence of arbovirus activity within a locale. Maternal antibodies may interfere with neutralizing responses in young foals. This response is confounded by recent vaccination. The most common neutralizing antibody test formats are the classic plaque reduction neutralization test (PRNT) and a microwell format test. Practical application of the microwell vs the PRNT indicates that the endpoint titer in the microwell test can be several logs higher than in the PRNT, so results cannot be compared between samples.
Several postmortem diagnostic assays are available and, although specific, they vary in sensitivity, depending on the virus. The midbrain and brain stem have the highest concentrations of encephalitic viruses, including rabies virus. Immunohistochemistry and PCR testing for EEEV and EHV-1 are relatively straightforward compared with that for WNV. Diagnostic testing continues to confirm the unreliability of detection of virus, even by PCR, because of the low viral load in WNV infection. Often, only a single neuron in one or two sections may yield positive virus staining in the horse by immunohistochemistry. When tested by PCR, the limited viral load dictates accurate testing of appropriate tissues consisting of several locations, including thalamus, hypothalamus, rostral colliculus, pons, medulla, and anatomically identified spinal cord. Viral isolation is still important for molecular epidemiology.
Infectious and noninfectious causes of brain and spinal cord diseases should be considered as differential diagnoses. Infectious causes include alphaviruses, rabies, equine protozoal myeloencephalitis (see Equine Protozoal Myeloencephalitis), and EHV-1; less likely causes are botulism and verminous meningoencephalomyelitis (eg, Halicephalobus gingivalis, Setaria spp, Strongylus vulgaris). Noninfectious causes include hypocalcemia, tremorigenic toxicities, hepatoencephalopathy, and leukoencephalomalacia.
Treatment:
Treatment of viral encephalitis is supportive, because there are no specific antiviral therapies. Management is focused on controlling pain and inflammation, preventing injuries associated with ataxia or recumbency, and providing supportive care. Intervention does not appear to significantly affect the outcome of most fulminate EEEV infections. For WNV, flunixin meglumine (1.1 mg/kg, IV, bid) early in the course of the disease decreases the severity of muscle tremors and fasciculations within a few hours of administration.
Recumbent horses that are mentally alert frequently thrash, causing self-inflicted wounds and posing a risk to personnel. Responses to tranquilizers and anticonvulsant medications are variable, depending on the virus and severity of disease. A sling and hoist may be used to assist horses that are recumbent and have difficulty rising; however, recumbent horses with EEEV generally are too comatose to sling. Dysphagic horses require fluid and nutritional support.
Until equine protozoal myeloencephalitis is excluded, prophylactic antiprotozoal medications may be instituted. Other supportive measures (eg, oral and parental fluids and nutrition for dehydrated and dysphagic horses) are also important. Broad-spectrum antibiotics should be given for treatment of wounds, cellulitis, and pneumonia. Horses with intermittent or focal neuropathies have a better prognosis than those with complete flaccid paralysis or that appear comatose. Efficacy of specific antiviral agents for treatment of naturally occurring WNV or EEEV infection is unknown, even in people. Recent work with passive immunotherapy indicates possible benefit after the onset of clinical signs in WNV models.
Prognosis:
Mortality of horses showing clinical signs from EEEV is 50%–90%, from WEEV 20%–50%, from VEEV 50%–75%, and from flavivirus infections 35%-45%. Horses with clinical neurologic signs from alphavirus infection that recover have a high incidence of residual neurologic deficits, whereas many horses that recover from WNV disease have been reported to have no residual neurologic deficits. In EEEV infection, death is frequently spontaneous. With WNV disease, horses are euthanized for humane reasons, but spontaneous death does occur. In EEEV, most surviving horses exhibit longterm neurologic signs. In WNV disease, overt clinical signs in horses that recover can last from 1 day to several weeks; improvement usually occurs within 7 days of onset of clinical signs. Although 80%–90% of owners report that the horse returns to normal function 1–6 mo after disease, at least 10% of owners report longterm deficits that limit athletic potential and resale value. Deficits include residual weakness or ataxia in one or more limbs, fatigue with exercise, focal or generalized muscle atrophy, and changes in personality and behavioral aberrations.
Prevention:
Vaccination against alphaviruses and flaviviruses as core annual vaccines are considered the standard of care for all horses in the USA as endorsed by the American Association of Equine Practitioners. Formalin-inactivated whole viral vaccines for EEEV, WEEV, and VEEV are commercially available in bi- and trivalent forms, usually formulated with tetanus toxoid; several now include WNV. Nonvaccinated adult horses require the label priming two injections. For adult horses in temperate climates, an annual vaccine within 1 mo before the start of the arbovirus season is recommended. However, for horses that travel between northern and southern areas affected by the virus, injections should be given two or even three times yearly in active arbovirus seasons. Mares should be vaccinated 1 mo before foaling to induce colostral antibody. If a pregnant mare is naive, the full priming series starting 2 mo before foaling is recommended. However, some mares do not produce colostral antibody if vaccinated for the first time during gestation.
In foals that have received adequate colostrum from vaccinated dams, vaccination should begin at 5–6 mo of age; foals should receive two additional vaccinations at 30 and 90 days after the first one. It is unclear whether maternal antibody interferes with vaccine responses in foals; however, epidemiologic evidence strongly indicates that horses between 4 mo and 4 yr old are highly susceptible to EEEV. If there is early spring (March) activity in the southeastern USA, horses may require three injections throughout the year, especially in horses <5 yr old and in horses that have recently arrived in the southeastern USA. In foals born to nonvaccinated or minimally vaccinated mares, maternal immunity may wane, and vaccination should be performed at 4, 5, and 6 mo of age. In Florida, where the highest numbers of EEEV cases occur, horses of all ages should be vaccinated in January and again in April before the peak of the season. A third vaccination should be administered late in the summer if the season is particularly active. The frequency of vaccination may be minimized to once or twice per year in climates that have short mosquito seasons and in which limited activity has historically been reported.
Currently, several whole inactivated virion vaccines, one recombinant inactivated vaccine, and a canarypox-vectored vaccine are licensed for prevention of WNV viremia in most of the Americas, including the USA and Canada, and Europe. An inactivated virus vaccine is readily available against Japanese encephalitis virus. There is experimental evidence that the canarypox WNV lineage 1 vaccine provides cross-protection against WNV lineage 2 infection. WNV challenge studies indicate that ~10% of properly immunized horses do not produce neutralizing antibodies to WNV and that 2.3%–3% of equine WNV cases seen in the field are in fully vaccinated horses. The duration of immunity from vaccination with the killed, adjuvanted WNV vaccine is unknown.
Protection of horses from arboviruses must also include efforts to minimize exposure to infected mosquitoes. Mosquito mitigation includes applying an insect repellent that contains permethrin on the horses at least daily during vector season, especially at times of day when mosquitoes may be most active. Environmental management is also essential and includes keeping the barn area, paddocks, and pastures cleared of weeds and organic material, such as feces, that might harbor adult mosquitoes. Cleaning water tanks and buckets at least weekly will reduce mosquito breeding areas. Removal of other containers such as flower pots and used tires that may hold stagnant water is essential for reducing the number of mosquitoes in the area.
Options for control of arbovirus infection in other animals emphasize reduction of exposure. Few arbovirus infections have been documented in dogs and cats; however, exposure to EEEV and WNV is detected in both species during arbovirus seasons. In active years, young dogs have been reported to be susceptible to EEEV infection.
Keeping dogs and cats indoors or in a screened area, especially during the time when mosquitoes are most active, reduces exposure. Disposal of dead birds or other small prey that might be eaten may reduce oral exposure.
Clinical cases of both of these diseases have also been recorded in other domestic and exotic animals during active seasons. Emus are exceptionally susceptible to EEEV. These animals are used for the commercial food industry, and infection results in high viremia and high amounts of virus shedding rectally, orally, and in regurgitated material. Vaccination will prevent viremia, shedding, and disease. Camelids are susceptible to WNV, and numerous reports of disease and pathology were recorded as the virus spread across North America. The killed, adjuvanted vaccine marketed for use in horses has been used in camelids without any reports of major adverse effects, and animals have demonstrated production of neutralizing antibody.
Zoonotic Risk:
The reporting of these diseases (or suspicion of such) on an international and national basis is a basic duty of all veterinarians to ensure the health and safety of horses and people. As such, a thorough diagnostic investigation to confirm these infections is imperative, irrespective of state resources available for subsidized testing.
People may be infected by most of the arboviruses that commonly cause viral encephalitis in horses. Clinical signs in people vary from mild flu-like symptoms to death. Children, the elderly, and those who are immunosuppressed are the most susceptible. People with neurologic disease due to arboviruses usually have permanent neurologic impairment after recovery. Human disease is reported infrequently and generally follows equine infections by ~2 wk. Veterinarians should be aware of the possibility of human infection and use repellents and other procedures to protect themselves from hematophagous insects when working in sylvatic virus habitats or handling viremic horses. In addition, waterproof clothing (gown, boots, gloves), respirator (N-95 or N-99 mask), face shield, and hair covering are recommended during all necropsies performed on horses suspected of having encephalomyelitis.
- Equine Arboviral Encephalomyelitis
- Overview of Equine Arboviral Encephalomyelitis
Also of Interest
Test your knowledge
Laryngeal paralysis is a common disorder of older, large-breed dogs. Denervation of the cricoarytenoideus dorsalis muscle of the larynx causes the vocal folds and arytenoid cartilages to fail to abduct during inspiration, creating an upper airway obstruction. Although the etiology is usually unknown, trauma and neoplasia can cause laryngeal paralysis. Which of the following conditions can also cause laryngeal paralysis?